
Biorobotics Laboratory BioRob
A Neural Visuomotor Controller for a Simulated Salamander Robot
Biljana Petreska - Master Thesis (February 2004)
 This work investigates the neural mechanisms underlying visually-guided behavior in salamanders, with a special attention to the interplay of neural processing and the biomechanics of the vertebrate body. We developed biologically motivated models of the retina, the optic tectum, the pretectum and the brain stem regions of the salamander brain, which were coupled to the spinal locomotor circuits and to the biomechanical model of a simulated salamander robot. The model integrates typical orienting, gaze stabilization, approaching and snapping behaviors specific to the salamander feeding activity, and also supports predation.
This work investigates the neural mechanisms underlying visually-guided behavior in salamanders, with a special attention to the interplay of neural processing and the biomechanics of the vertebrate body. We developed biologically motivated models of the retina, the optic tectum, the pretectum and the brain stem regions of the salamander brain, which were coupled to the spinal locomotor circuits and to the biomechanical model of a simulated salamander robot. The model integrates typical orienting, gaze stabilization, approaching and snapping behaviors specific to the salamander feeding activity, and also supports predation.
-
Diploma Thesis Report.
A complete literature review, all implemented models and respective results can be found in the diploma report.
Master Thesis Presentation.
The Optic Tectum Model
We show in a neuromechanical simulation how topological maps of neurons with fairly large receptive fields in the optic tectum can be used to reproduce salamander approach behavior to visual stimuli. With a simple structured mapping between the optic tectum and the brainstem, our salamander is capable of tracking a simple prey stimulus with high accuracy. The parameters of the linear activation function have been defined through an exhaustive search of the parameter space. We suggest that the sensorimotor transformation that occurs from the tectum to the brainstem is that of the horizontal angular distance of tectum neurons to amount of muscle activity (for a full description of the optic tectum model please refer to the corresponding chapter in the master thesis report). The processed visual input directly stimulates the neck motoneurons. Moreover the numbers of neurons used is biologically plausible. The decision to normalize the visual signal has made the network robust to all change in the stimulus parameters. We also suggest that even simpler models entail similar accuracy (i.e., with only contralateral input or leaving aside the offset parameters). Our model accounts for the characterictic saccadic pursuit movements, particular approach in monocularized animals and observed prey preferences.
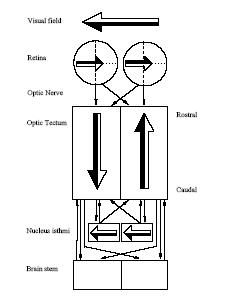
On the left. Schema of the visual information flow in the salamander brain. The tectal retinotopical representation of the visual field and direct connection between the optic tectum and the brain stem motor centers were the primary biological inspirations to the model investigated within this project.
-
Mapping of the tectum projections to the brainstem.
This figure also shows the representation of several stimuli in the tectum, which depends on the stimulus eccentricity. All points have identical elevation in the visual field as they are vertically aligned in the tectum.
First Tectum Model
In its first version the tectum model integrates tectal activity without normalizing it. Although neural weights can be deteremined to achieve excellent results with a particular type of stimulus, it is not robust to change in the stimulus parameters (i.e., size, contrast, etc..).
-
Illustration.
The salamander specilizes itself to a particular visual input (here a unique prey stimulus). With incremented tectal activity (distracting spheres) the behavior is unstable. NEW
Illustration (Mpeg-1, 770KB).
Illustration (DivX, 500KB)
Robustness of the Model
The salamander motoneurons are directly stimulated by integrated and normalized tectal activity. The salamander is able of accurately tracking a prey stimulus (i.e., orienting, gaze stabilization and approaching behavior). Our model is robust to any change of the stimulus parameters, such as stimulus size, speed, distance or contrast to the background (default parameters are a 16mm stimulus at 220mm distance and moving at 20°/s):
-
Stimulus speed of 4°/s.
Stimulus speed of 4°/s. (Mpeg-1, 8.4MB)
Stimulus speed of 4°/s. (DivX, 6.0MB)
Stimulus speed of 60°/s.
Stimulus speed of 60°/s. (Mpeg-1, 600KB)
Stimulus speed of 60°/s. (DivX, 370KB)
Stimulus size of 2mm.
Stimulus size of 2mm. (Mpeg-1, 1.4MB)
Stimulus size of 2mm. (DivX, 690KB)
Stimulus size of 80mm.
Stimulus size of 80mm. (Mpeg-1, 1.7MB)
Stimulus size of 80mm. (DivX, 1.1MB)
Stimulus distance of 15mm.
Stimulus distance of 15mm. (Mpeg-1, 1.4MB)
Stimulus distance of 15mm. (DivX, 910KB)
Stimulus distance beyond 800mm.
Stimulus distance beyond 800mm. (Mpeg-1, 1.4MB)
Stimulus distance beyond 800mm. (DivX, 860KB)
-
Optimal stimulus parameters for visually guided behavior in salamanders
used for the exploration of the weights function parameters.
Complex Background
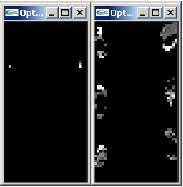 With a complex environment the salamander has some difficulties in following
prey stimuli, the amount of noise being considerable (see figure). This is not surprising at all as no selective or "winner-takes-all" mechanism was implemented. As can be observed from the simulations below, the salamander "sees" and orients his head toward the prey stimulus every time it is "more visible" (in front of a light background or stabilized salamander head). The animal also discriminates between objects with same apparent angular size (i.e., objects of different size situated at different distances from the salamander), however it can not be prevented from orienting its head toward "average flies".
With a complex environment the salamander has some difficulties in following
prey stimuli, the amount of noise being considerable (see figure). This is not surprising at all as no selective or "winner-takes-all" mechanism was implemented. As can be observed from the simulations below, the salamander "sees" and orients his head toward the prey stimulus every time it is "more visible" (in front of a light background or stabilized salamander head). The animal also discriminates between objects with same apparent angular size (i.e., objects of different size situated at different distances from the salamander), however it can not be prevented from orienting its head toward "average flies".
Integrating approach behavior is trivial with a unique stimulus. Ideally it should be coupled with a selective visual attention mechanism (i.e., enhanced retinal signals in the area containing the prey stimulus) and/or optokinetic or vestibucollic image stabilization reflexes (i.e., compensate for body undulations by antagonistic head movements) in order to cope with complex visual inputs (see example).
On the left. Difference between tectal activity with a simple stimulus and tectal activity with the same stimulus in a complex environment. Shows the amount of "noise" du to self-motion.
-
With distracting spheres in the background.
Same, DivX encoding, 1.1MB
Same, Mpeg-1 encoding, 1.7MB
Optic tectum neural network activity with distracting spheres in the background. TO COME
With two prey stimuli (one non-moving).
Same, DivX encoding, 1.1MB
Same, Mpeg-1 encoding, 1.7MB
Discriminating between objects of same apparent angular size.
Same, DivX encoding, 1.3MB
Same, Mpeg-1 encoding, 1.9MB
Following two coherently moving objects with distracting spheres.
Same, DivX encoding, 1.2MB
Same, Mpeg-1 encoding, 1.9MB
With a textured grass environment. TO COME
Prey-predator Discrimination
 Lesions of the pretectum facilitate feeding and abolish prey-predator discrimination (i.e., larger stimuli being interpreted as dangerous). Animals with unilateral lesions of pretectum respond with
feeding toward moving objects of any size and configuration (respond to everything
that moves including their own extremities and attack rather threatening
stimuli) within the visual field contralateral to the lesioned brain region and escape behavior failed to occur (third image). The size of the area in the visual field where feeding disinhibition occurs corresponds to the size of the lesions.
Lesions of the pretectum facilitate feeding and abolish prey-predator discrimination (i.e., larger stimuli being interpreted as dangerous). Animals with unilateral lesions of pretectum respond with
feeding toward moving objects of any size and configuration (respond to everything
that moves including their own extremities and attack rather threatening
stimuli) within the visual field contralateral to the lesioned brain region and escape behavior failed to occur (third image). The size of the area in the visual field where feeding disinhibition occurs corresponds to the size of the lesions.
We have implemented a simple prey-predator discrimination model which enables the salamander to follow prey and escape from predator stimuli (for a more detailed explanation see the diploma report). We have also proposed a model for tecto-pretectal interaction that reproduces the behavior of a salamander with pretectal lesions.
-
Prey Stimulus.
Prey Stimulus. (DivX, 190KB)
Prey Stimulus. (Mpeg-1, 630KB)
Predator Stimulus.
Predator Stimulus. (DivX, 700KB)
Predator Stimulus. (Mpeg-1, 1.4MB)
Salamander with Pretectal Lesions.
Salamander with Pretectal Lesions. (DivX, 460KB)
Salamander with Pretectal Lesions. (Mpeg-1, 970KB)
Monocularized Salamander
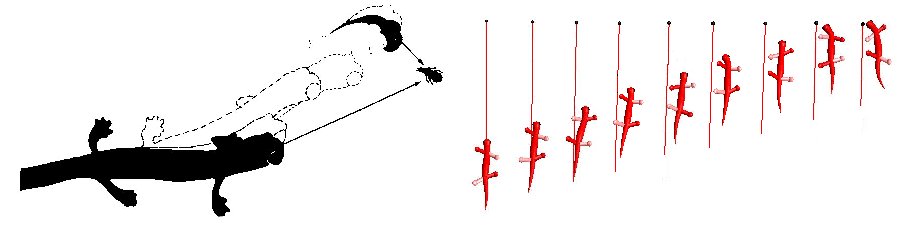
With one eye covered Hydromantes Italicus shows a conspicuous approach behavior toward a prey. It takes a curved path to approach the prey and bends his body away of it toward the side of the seeing eye, compensating the bending by turning the head between 60° and 90°. Our monocularized salamander exhibits the same approach behavior.
-
Movie of the monocularized salamander.
Movie of the monocularized salamander (DivX, 1.7MB)
Movie of the monocularized salamander (Mpeg-1, 2.4MB)
Prey Preferences
The experiment with double stimulus was reproduced. Just like real salamanders, our salamander (after a little "hesitation") prefers larger stimuli and preys of higher contrast. We insist on the fact that the salamander makes its decision upon its own sensory perception. In Simulander (previous work on the subject), the visual input to the tectum is a function of the stimulus size and the stimulus velocity (i.e., corresponds to the observer knowledge). We made our model contrast-sensitive by using a linear difference function (rather than binary) for the retinal ganglion cells of type 2. The salamander prefers larger prey objects since they enhance larger activation areas in the tectum. Faster objects are also preferred. They activate higher activity in the tectum (as they achieve larger eccentricity angles in less time). However for certain speeds, the salamander turns back to the slower stimulus, as suddenly it is projected to the peripheral visual field. This artefact would be completely suppressed by a visual attention mechanism. When two identical stimuli are presented the real salamander fails to respond in 45% of the cases (only 5% with different stimuli).
-
The salamander prefers objects of greater size.
The salamander prefers objects of greater size. (DivX, 380KB)
The salamander prefers objects of greater size. (Mpeg-1, 860KB)
The salamander prefers faster objects.
The salamander prefers faster objects. (DivX, 400KB)
The salamander prefers faster objects. (Mpeg-1, 1.0MB)
Objects with greater contrast elicit orienting behavior.
Objects with greater contrast elicit orienting behavior. (DivX, 360KB)
Objects with greater contrast elicit orienting behavior. (Mpeg-1, 880KB)
With identical stimuli the salamander can not decide.
With identical stimuli the salamander can not decide. (DivX, 250KB)
With identical stimuli the salamander can not decide. (Mpeg-1, 880KB)
B_fast_1.avi
B_fast_1_divx.avi
B_fast_1.mpg
Salamander Preferences. Movie of the double stimuli experiment with a real salamander, realized by Ursula Dicke.
Salamander Preferences. (DivX, 1.7MB)
Salamander Preferences. (Mpeg-1, 2.3MB)
-
The effect of stimulus features on the visual orienting behaviour of the salamander Plethodon jordani.
A study by Ursula Dicke.
 The double dummy experiment with a real salamander. By Ursula Dicke. The full study and movie are available above.
The double dummy experiment with a real salamander. By Ursula Dicke. The full study and movie are available above.
Snapping Model
A tongue was given to the salamander in order to investigate possible mappings between the optic tectum and brain stem that control the snapping behavior. A simple model with no new neurons as those used for the orienting behavior (i.e., using divergent projections), produces a coarse estimation of the stimulus distance and modulates the tongue protraction. The success rate is similar to that of S.salamandra (40%). Moreover our model predicts that lesions to the tectobulbar pathway result in overestimating the stimulus distance, which is consistent with real observations.
-
Snapping salamander with a frontally oriented head.
Snapping salamander with a frontally oriented head. (DivX, 710KB)
Snapping salamander with a frontally oriented head. (Mpeg-1, 4.8MB)
Snapping salamander. TO COME
Integrating approach and snapping behaviors. TO COME
 Some salamander species can protrude their tongue laterally and up to 80% of their body length (photo by Stephan M. Deban). Impressive movies of salamander feeding on the
Autodax Website.
Some salamander species can protrude their tongue laterally and up to 80% of their body length (photo by Stephan M. Deban). Impressive movies of salamander feeding on the
Autodax Website.
Relevant Topics
-
A 0.5mm stimulus at 50mm still elicits orienting behavior.
The visual acuity (minimal visible stimulus that elicits orienting behavior) has been falsely used to argument the need for an important number of tectal neurons (for more details see the section on number of tectal neurons in the diploma report).
(DivX, 760KB) Same film as above, DivX encoding
(Mpeg-1, 1.5MB) Same film as above, Mpeg-1 encoding
Experiments with irregularly moving stimulus.
(DivX, 480MB)
Same film as above, DivX encoding
(Mpeg-1, 1.5MB)
Same film as above, Mpeg-1 encoding
Following a stimulus with random speed.
(DivX, 2.0MB)
Same film as above, DivX encoding
(Mpeg-1, 2.9MB)
Same film as above, Mpeg-1 encoding
Links to Other Web Sites
- Archived student projects
- Alain Dysli
- Alexandre Tuleu
- Anurag Tripathi
- Ariane Pasquier
- Aïsha Hitz
- Barthélémy von Haller
- Benjamin Fankhauser
- Benoit Rat
- Bertrand Mesot
- Biljana Petreska
- Brian Jimenez
- Christian Lathion
- Christophe Richon
- Cédric Favre
- Daisy Lachat
- Daniel Marbach
- Daniel Marbach
- Elia Palme
- Elmar Dittrich
- Etienne Dysli
- Fabrizio Patuzzo
- Fritz Menzer
- Giorgio Brambilla
- Ivan Kviatkevitch
- Jean-Christophe Fillion-Robin
- Jean-Philippe Egger
- Jennifer Meinen
- Jesse van den Kieboom
- Jocelyne Lotfi
- Julia Jesse
- Julien Gagnet
- Julien Nicolas
- Julien Ruffin
- Jérôme Braure
- Jérôme Guerra
- Jérôme Maye
- Jérôme Maye
- Kevin Drapel & Cyril Jaquier
- Kevin Drapel & Cyril Jaquier
- Loïc Matthey
- Ludovic Righetti
- Lukas Benda
- Lukas Hohl
- Lukas Hohl
- Marc-Antoine Nüssli
- Martin Biehl
- Martin Riess
- Martin Rumo
- Mathieu Salzmann
- Matteo Thomas de Giacomi
- Matteo Thomas de Giacomi
- Michael Gerber
- Michel Ganguin
- Michel Yerly
- Mikaël Mayer
- Muhamed Mehmedinovic
- Neha Priyadarshini Garg
- Nicolas Delieutraz
- Panteleimon Zotos
- Pascal Cominoli
- Pascal Cominoli
- Patrick Amstutz
- Pedro Lopez Estepa
- Pierre-Arnaud Guyot
- Rafael Arco Arredondo
- Raphaël Haberer-Proust
- Rico Möckel
- Sacha Contantinescu
- Sandra Wieser
- Sarah Marthe
- Simon Blanchoud
- Simon Capern
- Simon Lépine
- Simon Ruffieux
- Simon Rutishauser
- Stephan Singh
- Stéphane Mojon
- Stéphane Mojon
- Sébastian Gay
- Vlad Trifa
- Yvan Bourquin